
Viville Jean-Sébastien
Le rôle des ions dans les facultés électriques de Gymnarchus niloticus


Introduction
Le groupe des poissons électriques se subdivise en deux
catégories principales : la première concerne les organismes qui
possèdent uniquement des électrorécepteurs. Ces poissons
perçoivent des champs électriques qu’ils n’ont pas
eux même engendré : il s’agit du groupe des électrosensoriels
passifs. Dans ce groupe on retrouve certaines espèces appartenant aux
familles des Squaliformes, des Siluriformes et des Rajiformes notamment. La
seconde catégorie regroupe les poissons qui sont munis en plus d’organes
électrorécepteurs, d’organes électrogènes
: il s’agit du groupes des électrosensoriels actifs. On y retrouve
des poissons électriques à force décharges plutôt
rares en eau douce telle que l’anguille électrique (Electrophorus
electricus). L’émission électrique intervient alors par
ravales occasionnelles et répond principalement à des fonctions
de défense et de prédation. D’autres espèces appartenant
à ce groupe sont dites faiblement électriques : leur faible décharge
électriques peuvent alors être soit pulsatoires soit ondulatoires,
cette seconde catégorie concerne des décharges électriques
de forme quasi sinusoïdales ne présentant aucune discontinuité
entre deux périodes successives, c’est le cas de l’espèce
étudiée ici : Gymnarchus niloticus, plus couramment appelé
le Gymnarque du Nil.
La question se pose alors de savoir quels sont les tissus responsables de l’électrogénèse
et de l’électroréception, les mécanismes mis en œuvre
et plus précisément le rôle des ions intervenants : Na +
et K+ . Nous verrons dans un premier temps une présentation du Gymnarque
du Nil, puis nous mettrons en évidence son pouvoir électrogène.
Nous verrons ensuite quelles sont les structures à l’origine de
cette caractéristique, ainsi que les propriétés électrorécéprtices
du poisson
I. Présentation du poisson
Gymnarchus niloticus semble être le premier poisson faiblement électrique
sur lequel des expériences décisives ont été faîtes
pour mettre en évidence l'existence et la fonction électroperceptive
des décharges autostimulatrices qu’il est capable d’émettre.
Cependant il est encore peu connu aujourd'hui, que ce soit du point de vue anatomique,
que du point de vue physiologique. Cela s'explique par une difficulté
technique d'expérimentation qui lui est tout à fait propre et
qui tient de sa constitution physique. En effet, sept fois par heure en moyenne
Gymnarchus niloticus doit, en complément de son insuffisante respiration
branchiale, monter à la surface pour renouveler sa provision d'air, sa
vessie gazeuse, spongieuse et fortement vascularisée jouant un rôle
pulmonaire. Aussi il est impossible de l’immobiliser sous l' eau pour
une durée supérieure à une dizaines, voire à une
vingtaine de minutes sans risquer de l' asphyxier, durée trop brève
pour pratiquer certaines expériences minutieuses et rigoureuses.
A. Famille :
Gymnarchus niloticus est le seul représentant de la famille des Gymnarchidae,
elle-même famille unique du sous-ordre des Gymnarchoidea, lui-même
inclus avec celui des Mormyroidae dans l'ordre des Mormyriformes. le genre Gymnarchus
se distingue de tous les autres représentants de cet ordre par bon nombre
de caractères : une vessie natatoire pulmonoïde et cellularisée,
un appendice caudal acuminé dépourvu de nageoire caudale, une
nageoire dorsale étendue sur toute la longueur du corps, un cerveau et
une oreille interne organisés différemment, l'absence de ceinture
pelvienne et de nageoire anale ...
Mormyridae et Gymnarchidae (c'est-à-dire Gymnarchus)
seraient issus les uns et les autres d’un ancêtre commun ayant vécu
au Jurassique. Il n'y a pas de rapport entre les Gymnotoïdes sud-américains
(qui sont des Ostariophysaires) et les Mormyriformes africains (qui sont des
Isospondyles), les continents américain et africain étant déjà
séparés quand les ancêtres des uns et des autres développèrent
les facultés d’électrogénèse, cependant le
Gymnarque , par la disposition de ses organes électriques, par son appendice
caudal acuminé et dépourvu de nageoire, par sa rigidité
posturale, par son mode de propulsion (ondulation réversible d'une nageoire
impaire unique) et par la forme de son onde électrique quasi-sinusoïdale
suggère un rapprochement avec les « Humbers », poissons originaires
d’Amérique du sud, ce qui constitue un exemple de convergence évolutive
intéressant.
B. Répartition géographique et
milieu de vie :
Gymnarchus niloticus est un prédateur qu'on ne rencontre qu'en Afrique,
dans une zone comprise entre le 5ième et 15ième degré de
latitude Nord, embrassant le cours et le bassin supérieur du Nil, le
bassin du Tchad, celui du Niger, du Sénégal, de la Gambie, il
est signalé dans le Lac Rodolphe.
Il habite les régions marécageuses, les zones d'inondation, et
vit dans des eaux sombres et limoneuses où la vue est peu efficace ou
totalement inopérante, d’où l’utilité de sa
« fonction » électrique.
C. Régime alimentaire et comportement :
Sa croissance est assez rapide, adulte il atteint et parfois dépasse
1.50 m. Son régime alimentaire est exclusivement carnivore : larves d’insectes,
crustacés, petits poissons, vers quand il est jeune ; gros poissons ou
batraciens à l'âge adulte. Il use parfois d'une « technique
» très remarquable pour capturer ses proies: il s'en approche silencieusement
(« au radar ») à quelques centimètres, puis brusquement
déclenche une aspiration violente, véritable implosion dont le
bruit est perceptible à. une certaine distance, transportant instantanément
la proie dans l’œsophage du prédateur.
Gymnarchus a un comportement territorial très prononcé en aquarium,
il ne supporte pas la présence d'un congénère. Dans la
nature il chasse vraisemblablement seul et de nuit, s'approchant sans bruit
ni remous par légères ondulations de la dorsale, de ses proies
préalablement détectées.
D. Reproduction :
Au mois de mai, dans les étangs couverts de nénuphars, les femelles
ont déjà l’ovaire rempli de gros œufs, en effet chez
tous les Mormyridés seule la gonade gauche se développe.
Les œufs sont sphériques, jaunes pâles et atteignent à
maturité un diamètre de 9-10 mm, taille tout à fait remarquable
pour des œufs de poisson. De juin à septembre, à la saison
des pluies, quand la crue du Niger a transformé la savane en marécages
et que reverdissent les graminées, c’est par 1,5 m à 2,20
m de fond que les gymnarques édifient leurs nids. A cette fin ils coupent
avec leurs dents des tiges de « Bourgou » (Echinochloa stagnina)
de 2 à 3 m de long, les replient et les entassent de manière à
former une masse flottante allongée de 1,5 m sur 0,8 m au centre de laquelle
a été aménagé une cavité de 50 cm sur 20
cm où sont déposés les œufs qui éclosent au
bout de 5 jours. Les juvéniles, une fois le sac vitellin résorbé,
quittent le nid vers le 18-ième jour.
Dans le nid les jeunes, entassés , font un incessant va et vient pour
happer l'air en surface ; ils ont déjà la nage des adultes et
leur remarquable aptitude à la marche arrière. Un comportement
protecteur de la part des parents est observé vis-à-vis de leur
progéniture, en effet les Gymnarques ont des ennemis sérieux :
les indigènes qui consomment leur chair et leurs œufs, et tes Polyptères
(Polyptèrus senegalus) qui pillent les nids. Dans les zones de reproduction,
les nids flottants sont en grand nombres, distants de 10 m à. peine.
Fin septembre tout est terminé et les nids sont abandonnés.
Gymnarchus niloticus est admirablement adapté à. la navigation
sans visibilité grâce à son sens électrique, c'est-à-dire
grâce à un système électrique de perception de l'environnement
qui lui permet de s’orienter dans l'espace, de localiser les obstacles,
de détecter des proies. Cette modalité sensorielle originale –
qui, on le sait depuis peu se rencontre chez quelque 500 espèces de Poissons
- présente deux aspects : l'un, actif, l’électrogénèse,
l'autre, passif ou du moins perceptif, l’électroréception.
La première permet un sondage électrique cadencé de l’environnement,
générateur d'informations ; la seconde réalise la collecte,
l’analyse et l'intégration de ces informations en une « image
» globale du milieu.
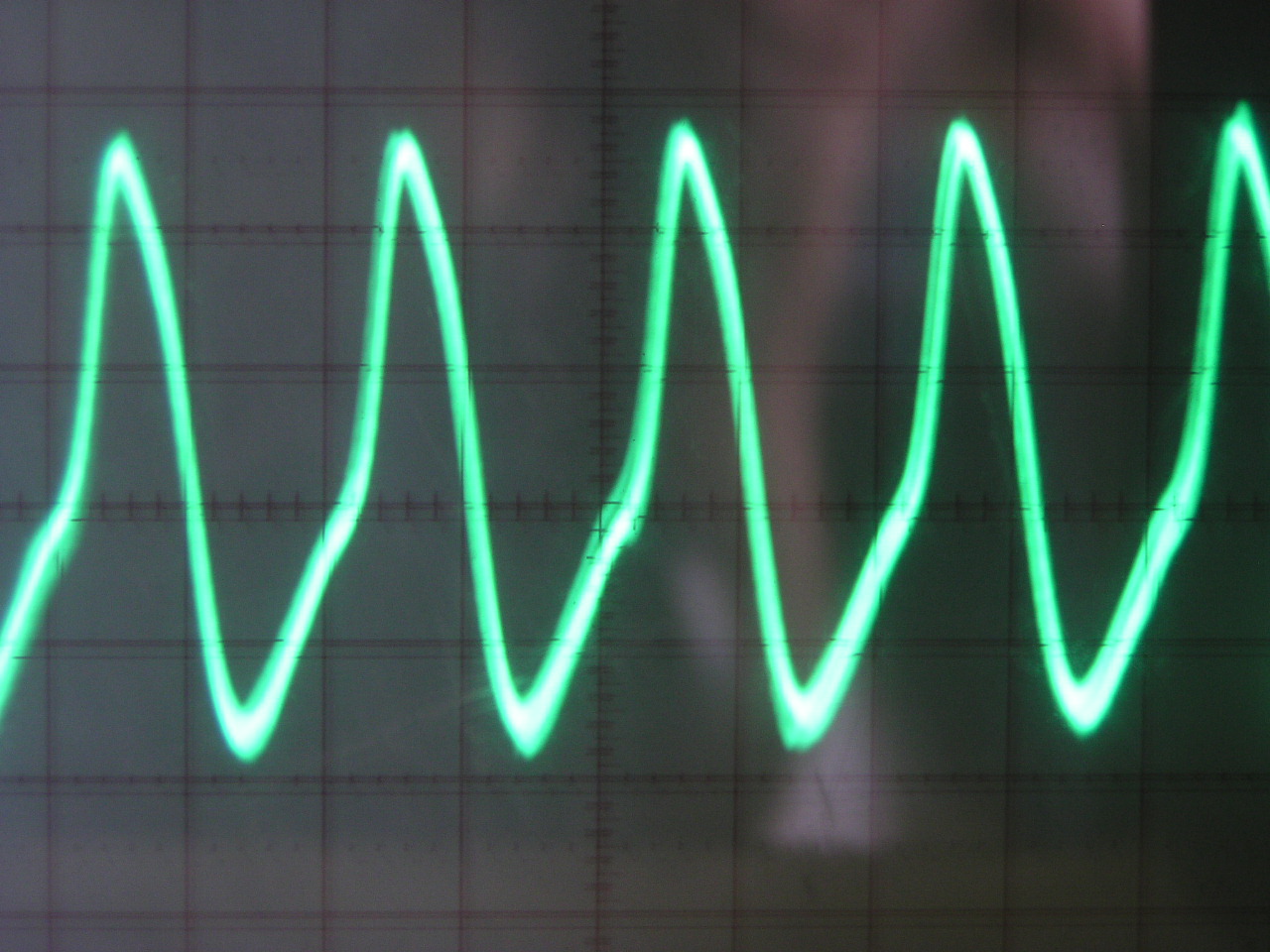
II. Expérience :
Cette expérience, destinée à mettre en évidence
l’extrême régularité de l’émission auto-stimulatrice
du Gymnarque du Nil. Elle fut présentée en première mondiale
par l’Aquarium tropical de Nancy à la Foire Exposition en juin
1983.
A. Montage et protocole
Deux électrodes verticales A et B en acier inoxydable plongent dans l’eau
de l’aquarium. A l’avant du bac l’électrode A et, à
l’arrière, l’électrode B déterminent un canal
de captage, qui recueille les variations ondulatoires du champ électrique,
dont le poisson s’entoure continuellement. Un amplificateur opérationnel
est nécessaire pour étudier le signal convenablement : l’impédance
d’entrée de l’amplificateur est de l’ordre de quelques
milliers d’ohms. Aussi, on a intérêt, pour recueillir convenablement
le signal, à couper toute composante continue éventuelles : mise
en série de résistances et condensateurs.
B. Résultats
On obtient un oscillogramme de type sinusoïdal, ce qui met en évidence
les variations périodiques de tensions ( de l’ordre de quelques
millivolts à quelques dizaines de millivolts) entre les deux électrodes.
On a donc deux phases : une phase d’ascension et une phase de chute du
signal (voir III.B). Ainsi, il y a bien électrogénèse.
III. Electrogenèse
A. Organisation générale
Le Gymnarque porte huit organes de forme tubulaires, de longueur inégale,
insérés par paires à l'arrière du poison, entre
les muscles de la queue, et se prolongeant vers l'avant un peu au-delà
de la moitié du corps. Ce sont les organes électrogènes.

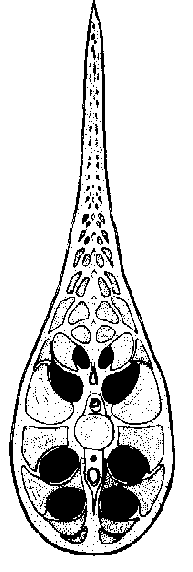
L'innervation en est assurée par huit faisceaux nerveux en provenance
de cellules de la corne ventrale, motrice, de la moelle épinière,
elles-mêmes sous la dépendance d'un centre situé dans la
formation réticulée du bulbe rachidien et qui constitue le «
pace-maker », le centre rythmeur de l'émission électrique.
Chacun est doté d'environ l40 cellules « appelées électroplaques
ou « électrocytes » génératrices de courant.
L’origine embryonnaire de ces cellules est myogénique (modification
évolutive de cellules musculaires) (il existe des familles de poissons
électriques (Apteronotidae) pour lesquelles ces cellules sont évolutivement
dérivées de fibres nerveuses).
B. Mécanisme
1. Première phase
D’un point de vue fonctionnel, chaque unité excitable est une cellule électrique limitée par une membrane, qui sépare deux milieux de composition chimique différente. La perméabilité sélective de cette membrane maintient activement une différence de concentration en ions entre le milieu extérieur, 10 à 15 fois plus riche en cations sodium Na+ que le milieu intérieur, qui lui est riche en anions organiques de grosse taille prisonniers de la membrane, et en ions potassium K+ qui, compte tenu de leur faible dimension diffusent à travers la membrane et vont enrichir sa charge positive externe ([K]int.= 30*[K]ext.). Cela se traduit par une différence de potentiel de quelques dizaines de millivolts, le potentiel de repos. Durant le phase de repos, le milieu extracellulaire est chargé positivement tandis que le milieu intracellulaire est chargé négativement.
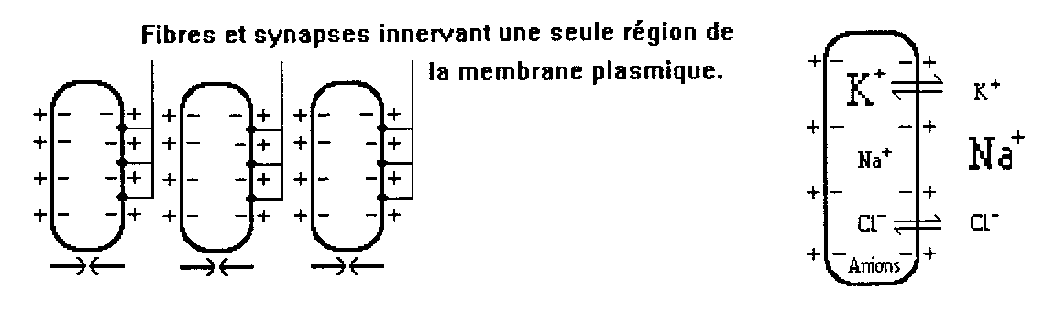
Sous l’effet de l’influx nerveux, qui est une onde de dépolarisation, la région innervée de la membrane entame sa dépolarisation. Elle devient pour un moment, à l’endroit de la stimulation, perméable aux ions Na +. Ces derniers profitent du gradient de concentration mais également du champ électrique pour faire éruption dans la cellule. Cela rend positive la face interne de cette région innervée et négative la face externe, du fait de la lacune en charges positives. Pendant ce temps la polarité de la région opposée (non innervée) de la membrane reste inchangée. Les deux faces opposées sont alors en série, telle une pile électrique. Il y a ainsi création d’un potentiel d’action, la décharge synchronisée de toutes les cellules engendrant un courant électrique dont le potentiel global est la somme des tensions cellulaires différentielles. Cette première phase traduit l’ascension du signal.
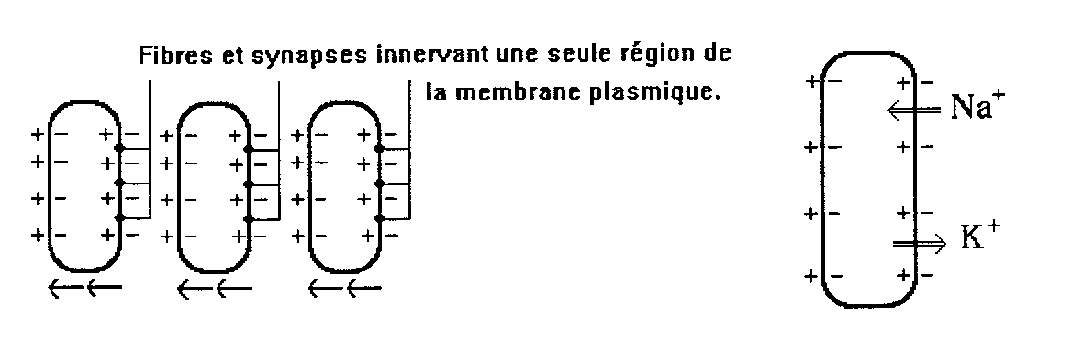

Electroplaques schématiques dont lequel on a figuré la répartition
des charges électriques sur la membrane soit dans l’état
de repos (à gauche) soit en moment du sommet de l’onde de décharge
(à droite). Les points noirs représentent les terminaisons nerveuses
sur l’électroplaque. On voit ce qui est essentiel : une seule face
est innervée. Au moment de l’activité, elle perd ses charges
(dépolarisation) par accroissement de perméabilité pour
les ions et même dans la plupart des cas, elles se polarisent pour un
instant en sens inverse, il se produit alors une addition des forces électromotrices
(60+80 = 140 mV) tandis qu’au repos elles se neutralisent (80-80=0).
2. Deuxième phase
Ensuite l’onde de dépolarisation atteint la face non innervée de la cellule, ce qui entraîne sa dépolarisation : elle devient positive du côté interne et négative du côté externe ; tandis que dans le même temps, la face innervée retrouve son état d’équilibre initial : positive du côté externe, négative interne. Comme lors de la première phase, les faces opposées sont en série, mais la tension change de signe, d’où la chute du signal, on a donc bien un courant alternatif.
Ce courant ondulatoire qui, à l’intérieur de l'organe, se dirige de l’arrière vers l'avant, referme son circuit de 1’ avant vers l arrière à travers l’eau ambiante, toujours assez ionisée, dans la nature, pour être conductrice. I1 se forme ainsi autour du Poisson un véritable champ électrique périodique.
3. Modalités du passage des ions
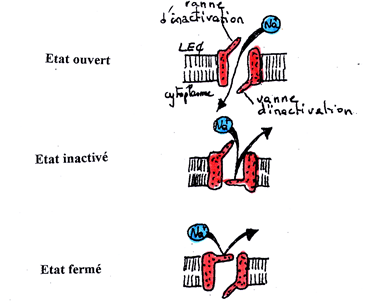
Initialement le passage des ions Na+ est bloqué en raison de l’état « fermé » des transporteurs, ce qui permet l’entretien du gradient électrique. Mais, lorsque le signal d’ouverture de ces transporteurs, qui est une onde de dépolarisation, arrive au niveau des transporteurs ioniques, ceux-ci s’ouvrent et entraînent le passage des ions, et donc l’émission d’un courant électrique. Ensuite, ces transporteurs se ferment progressivement au bout de 4 à 5 ms, ce qui entraîne la diminution progressive de l’entrée des ions Na+. L’état « fermé » de ces canaux, lorsque la différence de potentiel entre les deux membranes est nulle, est différent de l’état « fermé » initial. Ces transporteurs sont alors dans un état inactivé et ceci jusqu’à la repolarisation de la membrane, ils repassent alors dans un état fermé et le cycle recommence.
Les trois états conformationnels des canaux Na+ voltage dépendant.
Tout objet, inerte ou vivant, présentant une impédance électrique (résistance ohmique réactance capacitive etc...) différente de celle de l'eau environnante introduit une distorsion dans ce champ dont il allonge et disperse ou au contraire court-circuite et concentre les lignes de courant à proportion de sa mauvaise ou de sa. bonne conductibilité. Il s’en suit à. la surface de l'animal une modification locale de l'intensité du courant transcutané. C'est ici qu’interviennent les organes électrorécepteurs qui tapissent la peau et mesurent cette intensité .
IV. Electroréception
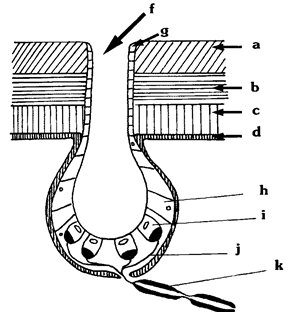
Vue au microscope la peau des Mormyriformes est un tissu épais et serré de cellules (souvent hexagonales) percées d'un grand nombre de pores qui sont les orifices des organes électrorécepteurs. Chez Gymnarchus ils pourraient atteindre le nombre d'une centaine de mille, répartis sur toute la surface du corps, mais avec une densité privilégiée sur la tête et la partie antérieure du tronc. De forme ampullaire ou tubéreuse, un récepteur mesure entre 60 et 250 microns. Logé sous la peau, il est mis en communication avec la surface par un canal à mucus ou une structure apicale jouant un rôle analogue. Sa cavité sensorielle proprement dite contient une gelée de polysaccharides. Gelée et structure intermédiaires sont beaucoup plus conductrices que la peau environnante. Les lignes de courant sont ainsi « focalisées comme par une lentille » et amenées à passer sélectivement par la cavité des organes électrorécepteurs, dont le fond est tapissé de cellules sensorielles. Gymnarchus possède au moins trois sortes d'électrorécepteurs dont les deux principaux sont les organes ampullaires et les tubéreux (au sens large ).
A. Les organes ampullaires (Micloampullae).
Ils permettent au poisson d’évaluer la distance qui le sépare
des éléments qui l’environnent. Leur fonctionnement est
asynchrone, c'est-à-dire indépendant de l'activité des
organes électrogènes du Poisson. Ils sont surtout sensibles aux
champs électriques continus ou de basse fréquence, fussent-il
extrêmement faibles, tels qu'en produisent sans exception tous les êtres
vivants et en particulier tous les poissons, même dépourvus d'organes
électrogènes spécialisés. Les fibres nerveuses afférentes,
issues des organes ampullaires acheminent vers les centres nerveux supérieurs
l’information relative à l'intensité des champs détectés.
Cette information est codée ici en impulsions nerveuses modulées
en fréquence dans sa fibre affluente une information codée.
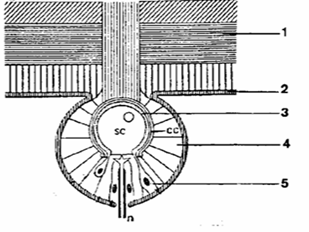
B. Les organes tubéreux
La portée effective de ces organes est supérieure à celle
des organes ampullaires (7 à 8 mètres environ), ils apportent
à l’animal des renseignements quant à la nature des éléments
qui l’entourent sans pour autant lui fournir une information de distance.
Ils forment une capsule épidermique contenant des cellules sensorielles,
mais sans communication directe avec l’extérieur. Situés
dans une invagination de la membrane basale de l’épiderme, ils
sont nettement moins sensibles que les électrorécepteurs ampullaires.
Cette structure sensorielle n’est différenciée que chez
les espèces électrogènes à faibles décharges
et ont deux rôles principaux : l’électrolocalisation et l’électrocommunication.
Ces organes tubéreux sont sensibles à l’intensité
du champ électrique fourni par l’émission électrique
cadencée du poisson et décèlent les modifications que lui
font subir les objets extérieurs du fait de leur conductivités
hétérogènes. Les différentes valeurs d’intensité
recueillies par ces électrorécepteurs sont exprimées en
impulsions modulées en fréquences dans la fibre afférente
et acheminées jusqu’au SNC. Le seuil de sensibilité des
organes tubéreux varie périodiquement : variation synchrone avec
l’émission électrique de sondage :minimum au moment de l’émission-réception
d’une impulsion de sondage, le seuil s’élève ensuite
(protection contre le brouillage) pour s’abaisser à nouveau au
moment de l’impulsion suivante .
L’électrolocalisation chez Gymnarchus niloticus consiste en un
courant ondulatoire qui, à l’intérieur de l’organe,
se dirige de l’arrière vers l’avant, referme son circuit
de l’avant vers l’arrière à travers l’eau ambiante.
Il se forme autour du poisson un véritable champ électrique périodique.
Tout objet, inerte ou vivant, présentant une impédance électrique
différente de celle du poisson produit une distorsion dans ce champ (allongement
ou au contraire concentration des lignes de courant, respectivement suivant
sa mauvaise ou bonne conductivité). Il s’ensuit à la surface
de l’animal une modification locale de l’intensité du courant
transcutané. « L’ombre électrique » des objets
environnants se projette en deux dimensions sur la peau du poisson, véritable
rétine électrique.
Conclusion
Ainsi le Gymarque du Nil est une des nombreuses espèces
électrogènes que l’on peut rencontrer chez les Poissons.
En effet il est capable de produire un signal électrique afin de s’orienter
dans le milieu dans lequel il évolue, qui est souvent trouble. Cette
faculté remarquable permet de combler ses lacunes visuelles, peut être
apparues au cours de l’évolution en raison de son milieu de vie.
L’émission électrique produite joue uniquement un rôle
de « sonar » contrairement à d’autres espèces
qui utilisent leurs facultés électrogènes dans un rôle
de prédation, telle que l’anguille électrique.
Les ions impliqués dans ce processus sont donc le Sodium et le Potassium
principalement. Ils sont à l’origine du signal électrique
lui-même entraîné par un signal de dépolarisation
des organes concernés, en provenance du cerveau.
Les modalités de propagation de ce signal sont complexes et difficiles
à étudier, ce qui explique le peu de recherches effectuées
sur ce poisson à ce jour, alors que d’autres poissons électrogènes
sont beaucoup plus étudiés et utilisés par l’Homme
: cas des bioréacteurs et de la détection de la pollution de l’eau
(avec notamment Apteronotus Albifrons).
Ces facultés d’utilisation d’un champ électromagnétique
sont intéressantes, il semblerait qu’il existe d’autres êtres
vivants capables de se repérer grâce au champ magnétique
terrestre : les oiseaux migrateurs (?). Aussi, même chez l’Homme,
on constate que certains individus présentent des facultés magnétiques
particulières (sourciers).
Bibliographie
Florion, A., 1984, Essai d’utilisation de Gymnarchus
niloticus Cuvier 1829 pour la régulation d’un biochronomètre
à l’Aquarium tropical de Nancy, issu de Revue française
d’aquariologie.
Dautel, O., 2002, Les cellules au sein de l’organisme : la communication
intercellulaire chez les organismes animaux.
Thomas, M., 1995, Thèse sur la détection de la pollution de l’eau.
Remerciements
Nous tenons à remercier l’aquarium de Nancy, et plus particulièrement
M. Florion pour sa précieuse aide et sa générosité,
ainsi que Pauline Cochet.